Human cytomegalovirus (HCMV) is an extremely common member of the herpesvirus family. It is generally harmless, producing no symptoms, and most people become infected with HCMV at some point in their lives. While it may seem innocuous enough to the general population, this is far from the case; it is also the most prevalent cause of congenital diseases and severe birth defects in newborns, which often leads to death. It can also opportunistically infect patients with compromised immune systems, also leading to near-certain death. Researchers have thus spent years developing potential vaccines for HCMV, but a robust vaccine has not yet been introduced on a large scale. However, the future of HCMV vaccination is hopeful - researchers Tomic et al. have engineered a strain of HCMV which is capable of stimulating both our innate and adaptive immune response to the virus.
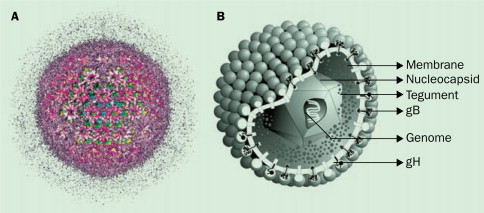
Human Cytomegalovirus Particle
So how exactly Tomic et al. engineer a version of HCMV which would induce a stronger immune response against itself rather than try to hide from our immune system as it usually does? They started with a cloned strain of HCMV known as TB40, which is capable of replicating at a high rate but does not code for many of its immunoevasins, proteins which help it evade our immune system. Next, they deleted one of the remaining immunoevasins, UL16, from the viral genome of TB40 and and inserted the ULBP2 gene in its place. The protein created by the ULBP2 gene, which is also called ULBP2, manifests on the surface of infected cells and binds to a receptor called NKG2D on Natural Killer (NK) cells, immune cells which protect our bodies against viral infection. When ULBP2 binds to NKG2D it heightens the NK cell’s regulatory capability, bolstering its ability to destroy cells infected with HCMV and other viruses.
The first major test ran by Tomic et al. was a NK cell cytotoxicity test, a test to determine how the mutant virus altered NK cells’ ability to destroy infected cells. They infected human foreskin fibroblast (HFF) cells with the ULBP2-TB40 mutant, introduced NK cells, and measured the percentage of mutant-infected cells which were destroyed, comparing this to the results of the standard TB40 strain. They discovered that a significantly higher percentage of mutant-infected cells were destroyed, indicating the success of the mutation in increasing the regulatory capabilities of NK cells.
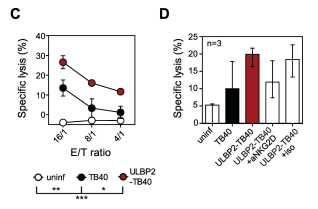
Fig. 1c The cytotoxicity test revealed that a significantly higher portion of ULBP2-TB40 mutant virus was destroyed when compared to standard TB40 virus. The mutant was successful in increasing the ability of NK cells to regulate the virus.
After determining the success of the ULBP2-TB40 mutant strain in cell culture, Tomic et al. made the major step to examine its effects in vivo - that is, in a living organism. They selected mice which had human hematopoietic stem cells, because these mice have been shown to model HCMV infection. They infected these “humanized” mice with ULBP2-TB40 to examine how their immune systems responded to the presence of the mutant virus, and compared these results to humanized mice infected with the standard TB40 strain. The quantity of viral DNA, as well as the quantity of CD8+ T cells (adaptive immune system cells) specific to cytomegalovirus, was then measured in the spleens of sacrificed mice. There was lower quantity of viral DNA in mice infected with the ULBP2 mutant when compared to standard TB40; this was likely related to the presence of human NK cells in the sacrificed mice; the ULBP2 mutation allowed the NK cells destroy more infected host cells. Compared to TB40-injected mice, ULBP2-TB40 mice also showed similar levels of CD8+ T cells specific to cytomegalovirus. This suggested that the mutant form of HCMV was still capable of stimulating our adaptive immune system despite the alterations made to its genome. This was important because having T cells specific to HCMV present before potential infection allows the body to more quickly neutralize its effects.
Next, Tomic et al. considered the fact that HCMV is a well-known inhibitor of the maturation of dendritic cells, which stimulate the production of T-cells. They knew that the TB40 strain of HCMV allows dendritic cells to mature and stimulate T-cell production, and wanted to ensure that expression of the ULBP2 protein would not hinder this process. They infected dendritic cells with TB40 and ULBP2-TB40, and used ex-vivo expansion to examine how effectively these dendritic cells activated different immune cells. Promising results were obtained; there was no notable difference in percentage of induced immune cells when compared to values from standard TB40 (Fig. 3a).
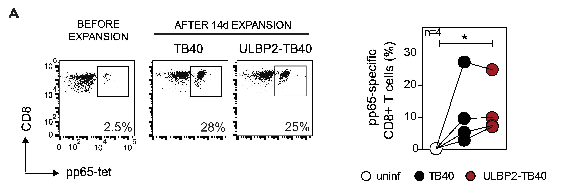
Fig. 3a Results of ex-vivo expansion show that HCMV-specific T cells are produced to a similar percentage when ex vivo expansion is conducted with TB40 and ULBP2-TB40-infected dendritic cells.
Tomic et al. further analyzed the maturation of dendritic cells infected with ULBP2-TB40 and discovered that these cells had a significant decrease in expression of MHC class I, a protein which can interact with T-cells to trigger an immune response. In fact, the ULBP2-TB40 mutant decreased the presence of MHC class I protein on cells to the same degree as a strain of HCMV which contains all of its immunoevasins. This is surprising in particular because MHC class I proteins activate virus-specific T cells, yet their significant decrease in the presence of ULBP2-TB40 did not lower the quantity of activated T cells.
This led Tomic et al. to the final stage of their research, trying to better understand the link between presence of the ULBP2 protein and activation of CD8+ T cells. They discovered that the presence of NKG2D - the receptor on NK cells which binds to ULBP2 to increase NK regulatory function - was significantly lowered on HCMV-specific T cells when they were activated by dendritic cells infected with ULBP2-TB40. However, the T cells were activated at a similar percentage regardless of which infected dendritic cells they were cultured with. This suggests that HCMV-specific T cells are activated by a different process when in the presence of ULBP2-TB40 infected cells (as opposed to TB40 cells), and this process likely has something to do with decrease of expression of NKG2D on their surface.
. Such a study shows how difficult it is to develop an antiviral vaccine; unpredictable interactions between viral proteins and cell receptors, inexplicable test results, and unforseen complications introduced by simple experiments are just a few of the challenges facing researchers. However this study also provides a hopeful outlook for the development of an HCMV vaccine. If researchers can better how ULBP2 downregulates surface content of NKG2D in HCMV-specific T-cells, and how NKG2D interacts with MHC class I within these T cells, they may be able to refine their mutant and take another step toward the development of a competent vaccine.
Article
Tomić A, Varanasi PR, Golemac M, Malić S, Riese P, Borst EM, et al. (2016) Activation of Innate and Adaptive Immunity by a Recombinant Human Cytomegalovirus Strain Expressing an NKG2D Ligand. PLoS Pathog 12(12): e1006015. doi:10.1371/journal.ppat.1006015
Additional References (found in hyperlinks)
Cheeran, Maxim C.-J., James R. Lokensgard, and Mark R. Schleiss. “Neuropathogenesis of Congenital Cytomegalovirus Infection: Disease Mechanisms and Prospects for Intervention.” Clinical Microbiology Reviews 22.1 (2009): 99–126. PMC. Web. 11 Dec. 2016.
Riss TL, Moravec RA, Niles AL. Cytotoxicity testing: measuring viable cells, dead cells, and detecting mechanism of cell death. Methods Mol Biol. 2011;740:103–14. doi: 10.1007/978-1-61779-108-6_12.
Eaves, Connie J.. "Hematopoietic stem cells: concepts, definitions, and the new reality." Blood 125.17 (2015): 2605-2613. Web. 10 Dec. 2016.
Brehm, Michael A., Leonard D. Shultz, and Dale L. Greiner. “Humanized Mouse Models to Study Human Diseases.” Current opinion in endocrinology, diabetes, and obesity 17.2 (2010): 120–125. PMC. Web. 11 Dec. 2016.
Harty, John T., Amy R. Tvinnereim, and Douglas W. White. "CD8+ T Cell Effector Mechanisms In Resistance To Infection." Annual Review Of Immunology 18.1 (2000): 275. Academic Search Premier. Web. 11 Dec. 2016.
Banchereau J. & Steinman R. M.. Dendritic cells and the control of immunity. Nature 392, 245–252 (1998).
Image credits
- Figure 1c from article.
- Figure 3a from article.
{kind=link}
No comments:
Post a Comment